








細胞自噬在維持健康和治療疾病中的應用
 供稿:市場部
供稿:市場部 發布時間:2023-07-25
發布時間:2023-07-25 瀏覽量:791次
瀏覽量:791次
Foreword
自噬是一種細胞內部的機制,通過將細胞內成分包裹起來并運輸到溶酶體進行降解,實現細胞的自我調節和清除有害物質。1963年,Christian de Duve首次提出了自噬的概念。1990年,Yoshinori Ohsumi通過釀酒酵母實驗直觀展現了自噬現象,并揭示了酵母和人的細胞中的自噬機制。2003年,自噬關鍵基因被命名為ATG基因,相關研究逐漸增加。2016年,Yoshinori Ohsumi獲得諾貝爾醫學或生理學獎,自噬研究進一步受到關注,成為生物學、醫學、植物學和微生物學的熱門研究領域。自噬的發現對于細胞調節和藥物開發具有重要意義。

巨自噬(macroautophagy):通過形成具有雙層膜結構的自噬體包裹胞內物質,最終自噬體與溶酶體融合。
微自噬(microautophagy):通過溶酶體或液泡表面的形狀直接吞沒特定的細胞器。
分子伴侶介導的自噬(chaperone-mediated autophagy,CMA):具有KEFRQ樣基序的蛋白在HSP70伴侶的幫助下,通過LAMP-2A轉運體轉運到溶酶體。
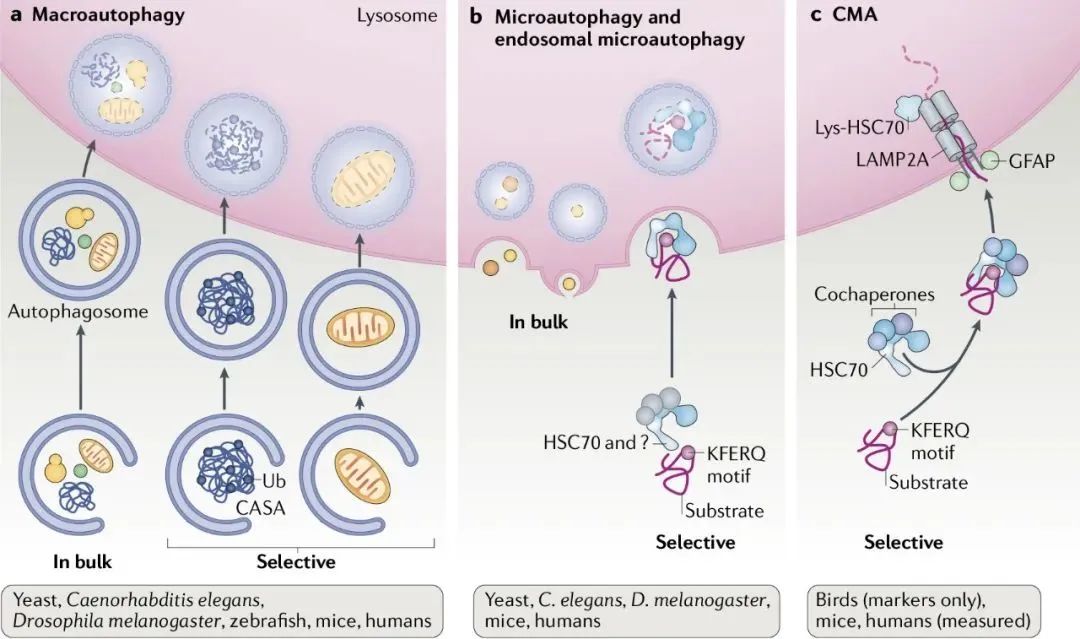
圖源[1]:https://www.nature.com/articles/s41580-018-0001-6
自噬可防止細胞損傷,促進細胞在營養缺乏的情況下存活,并對細胞毒性刺激作出反應。自噬包括生理條件下的基礎型自噬和應激條件下的誘導型自噬。基礎自噬是一種在大多數細胞中持續發生而水平相對較低的細胞自噬過程,對細胞內物質的更新及內穩態的維持具有不可或缺作用;誘導自噬則屬于一種應激狀態,其發生程度明顯強烈,在細胞應對外界惡劣環境維持生存中發揮重要功能。
自噬過程及核心蛋白
自噬過程主要分為自噬誘導階段、前體成核階段、延伸階段、自噬體成熟階段及自噬體與溶酶體融合階段。
01 自噬誘導階段(Induction)
02 前體成核階段(Nucleation)
03 延伸階段(Elongation)
自噬發生過程中有兩組類泛素化修飾過程, 分別發生在Atg5-Atg12-Atg16連接系統和Atg8/LC3連接系統中, 用于隔離膜的延長和自噬泡的形成。
在Atg5-Atg12-Atg16連接系統中,Atg12首先由類E1泛素活化酶Atg7活化。Atg12傳遞給類E2泛素轉移酶Atg10,與Atg10形成硫酯鍵。最后,Atg12與Atg5共價結合,形成Atg12-Atg5復合物。這個復合物在自噬發生時與Atg16結合形成Atg12-Atg5-Atg16。
LC3是酵母Atg8分子在哺乳動物中的同源物之一, Atg8的其他同源物還有GABARAP和GATE-16等。Atg8同源分子相比, LC3是被用作自噬發生的一個標志蛋白。LC3合成后,被AGT4B切割成LC3-I,自噬發生時,LC3-I被ATG7活化并與其形成硫酯鍵,然后傳遞給ATG3,最終在ATG5-ATG12-ATG16復合物的作用下,形成具有膜結合能力的LC3-II。參與自噬體的銜接。
04 自噬體成熟階段與自噬體與溶酶體融合階段
自噬體擴張和封閉過程包括自噬體外膜上ATG蛋白的清除,招募溶酶體傳遞蛋白和介導融合的蛋白。自噬體與溶酶體融合形成自噬溶酶體,自噬體內膜和內容物被溶酶體內的脂酶和蛋白酶降解為小分子,再利用。細胞骨架成分、運動蛋白、栓系因子、磷脂和SNAREs復合物在確保融合過程中起關鍵作用。
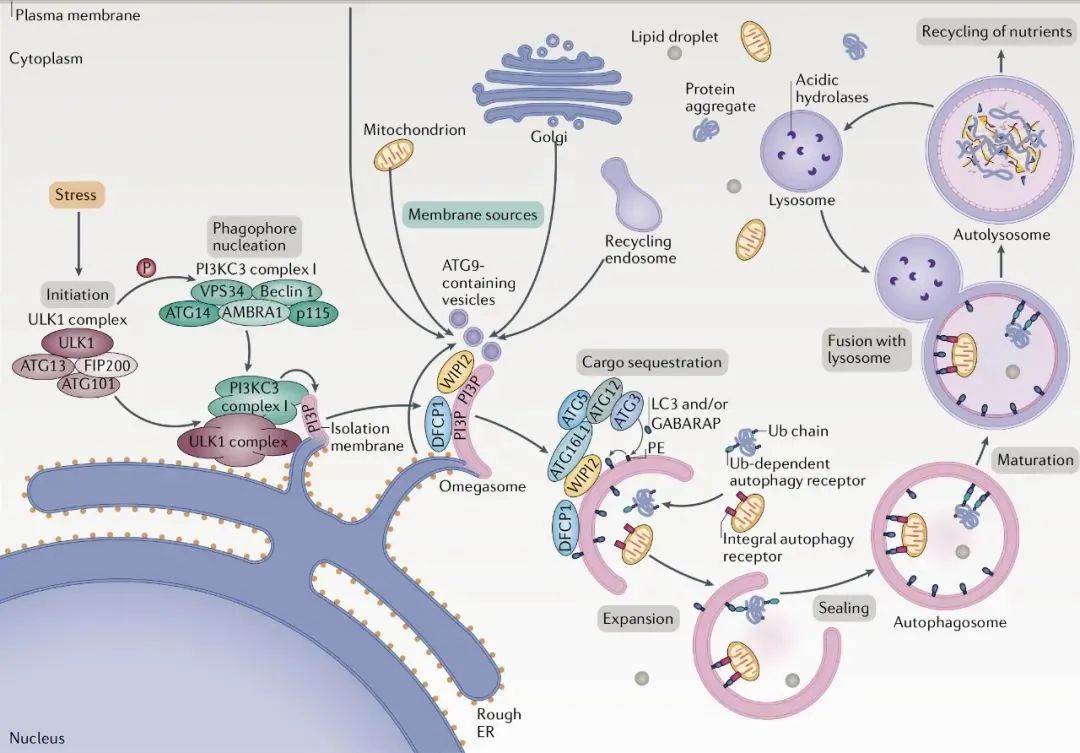
以下主要闡述起始及吞噬泡成核階段和細胞吞噬泡延伸階段參與的核心蛋白
1 ULK1蛋白激酶復合體中的ATG1,其功能是絲氨酸/蘇氨酸激酶;通過自噬機制的磷酸化組分啟動自噬;被AMPK或者mTORC1磷酸化。
2 VPS34:其功能是促進行P13KC3-C1復合體的形成,ULK1人磷酸微位點,穩定ULK1復合物;
3 Atg6/Beclin1:其功能是促進PI3KC-C1復合體的形成,調節脂質激酶VPS34;ULK1或AMPK磷酸化位點,促進自噬。
4 Atg9/mAtg9:Atg9是迄今發現的唯一一個編碼跨膜蛋白的Atg基因, 可能通過影響膜泡運輸對自噬發生起調控作用,其功能是將膜蛋白組分輸送到吞噬泡。
5 Atg5-Atg12-Atg16:ATG5直接結合膜,這種膜結合受ATG12的負調節,但被ATG16激活。ATG12-ATG5-ATG16復合物的膜結合是有效促進ATG8脂化(LC3-I轉化為LC3-II)所必需的。
6 ATG8/LC3:以LC3-Ⅰ和LC3-Ⅱ兩種形式存在;參與自噬體膜的形成,與自噬體膜表面的PE結合,可作為自噬體的標記分子。
自噬與疾病研究
許多疾病,諸如癌癥、神經系統疾病、炎癥與免疫反應、發育和衰老等都與細胞自噬密切相關。科學研究者也要積極探索自噬與這些疾病之間的聯系,努力揭示自噬調控分子機制,通過抑制或促進調節自噬過程,從而為各種疾病治療提供新的方向和可能性。
自噬與神經系統疾病
臨床研究發現,在許多神經退行性疾病中, 病變區域常有大量的泛素化蛋白質聚合物,這些多因錯誤折疊而被泛素化的蛋白質聚合物正是引起神經退行性疾病的重要原因之一。
小鼠中特異性敲除神經元中的自噬基因ATG5或ATG7,會抑制自噬發生的水平,同時伴隨著大量泛素化蛋白質聚合物的積累,并最終引發神經退行性疾病[3-4]。在小鼠和果蠅模型中,用Rapamycin或其類似物抑制mTOR的活性促進自噬發生,可以緩解神經退行性疾病的癥狀[5-6]。
由此可見,誘導自噬可能成為一種治療神經系統疾病的治療方法。其次是包括帕金森癥在內的一些神經退行性疾病。
自噬與腫瘤發生的聯系是當今自噬研究的一個重要熱點問題。自噬功能正常, 對腫瘤起抑制作用, 而敲除自噬基因將引發腫瘤的形成。Beclin1其實就是一個腫瘤抑制基因,干擾Beclin1的表達使自噬受到抑制, 會增加腫瘤發生的可能[7]。在胸腺癌細胞系中過表達Beclin1, 可以減緩癌細胞的生長速度并減少其致癌性[8]。UVRAG能夠結合Beclin1/Vps34復合物促進自噬, 而UVRAG同時有抑制腫瘤的功能[9]。Bif-1可以通過UVRAG結合Beclin1增強自噬, 敲除Bif-1, 顯著加快了腫瘤的生長[10]。
自噬受阻可能導致細胞失去生長調節及啟動程序性死亡的能力, 同時還可能阻礙細胞分解代謝, 使p62、受損的線粒體、蛋白質聚合物、ROS等有害物質積累, 并進一步損傷基因組DNA的穩定性, 致使原癌基因激活, 而以上這些最終都將導致腫瘤的發生。
因此,在不同的環境因素或穩定狀態下,自噬既可以阻止也可以促進腫瘤的發生。
自噬與免疫
自噬是至今已發現的唯一可以降解細胞器和較大蛋白質聚集物的細胞降解過程,因此也被預測可能參與降解外來微生物的過程[11]。許多研究發現,一些自噬蛋白甚至整個自噬過程都直接參與免疫和炎癥反應[12]。巨噬細胞中,干擾素IFN-γ可以誘導自噬來抵御分枝桿菌等病原菌,而且在已感染的巨噬細胞中, 用IFN-γ預處理可以引起自噬泡吞噬含有病原菌的膜泡并與溶酶體融合,增強其對胞內細菌的殺傷力。血管平滑肌細胞中, 腫瘤壞死因子TNF-α通過激活JNK通路和抑制Akt,上調LC3和Beclin1的表達促進自噬[13]。
由此可見,自噬與細胞免疫的發生必然存在密切的聯系。
【參考文獻】
1.Kaushik S, Cuervo AM. The coming of age of chaperone-mediated autophagy. Nat Rev Mol Cell Biol. 2018 Jun;19(6):365-381.
2.Dikic I, Elazar Z. Mechanism and medical implications of mammalian autophagy. Nat Rev Mol Cell Biol. 2018 Jun;19(6):349-364.
3.Hara T, Nakamura K, Matsui M, Yamamoto A, Nakahara Y, Suzuki-Migishima R, et al. Suppression of basal autophagy in neural cells causes neurodegenerative disease in mice. Nature 2006; 441(7095): 885-9.
4.Komatsu M, Waguri S, Chiba T, Murata S, Iwata J, Tanida I, etal. Loss of autophagy in the central nervous system causes neurodegeneration in mice. Nature 2006; 441(7095): 880-4.
5.Sarkar S, Perlstein EO, Imarisio S, Pineau S, Cordenier A, Maglathlin RL, et al. Small molecules enhance autophagy and reduce toxicity in Huntington’s disease models. Nat Chem Biol 2007; 3(6): 331-8.
6.Ravikumar B, Vacher C, Berger Z, Davies JE, Luo S, Oroz LG, et al. Inhibition of mTOR induces autophagy and reduces toxicity of polyglutamine expansions in fly and mouse models of.
7.Yue Z, Jin S, Yang C, Levine AJ, Heintz N. Beclin 1, an autophagy gene essential for early embryonic development, is a haploinsufficient tumor suppressor. Proc Natl Acad Sci USA 2003; 100(25): 15077-82.
8.Liang XH, Jackson S, Seaman M, Brown K, Kempkes B, Hibshoosh H, et al. Induction of autophagy and inhibition of tumorigenesis by beclin 1. Nature 1999; 402(6762): 672-6.
9.Liang C, Feng P, Ku B, Dotan I, Canaani D, Oh BH, et al. Autophagic and tumour suppressor activity of a novel Beclin1-binding protein UVRAG. Nat Cell Biol 2006; 8(7): 688-99.
10.Takahashi Y, Coppola D, Matsushita N, Cualing HD, Sun M, Sato Y, et al. Bif-1 interacts with Beclin 1 through UVRAG and regulates autophagy and tumorigenesis. Nat Cell Biol 2007; 9(10): 1142-51.
11.Kraft C, Peter M, Hofmann K. Selective autophagy: Ubiquitinmediated recognition and beyond. Nat Cell Biol 2010; 12(9): 836-41.
12.Jia G, Cheng G, Gangahar DM, Agrawal DK. Insulin-like growth factor-1 and TNF-alpha regulate autophagy through c-jun N-terminal kinase and Akt pathways in human atherosclerotic vascular smooth cells. Immunol Cell Biol 2006; 84(5): 448-54.
13.Gutierrez MG, Master SS, Singh SB, Taylor GA, Colombo MI, Deretic V. Autophagy is a defense mechanism inhibiting BCG and Mycobacterium tuberculosis survival in infected macrophages.Cell 2004; 119(6): 753-66

 返回列表
返回列表
在線咨詢
Online consultation
-
 在線咨詢
在線咨詢
-
 技術支持
技術支持

關注微信公眾號

